BIO 554/754
Ornithology
Avian Reproduction:
Anatomy & the Bird Egg |
|
Reproductive Anatomy:
- Gonads - paired testes in males & usually a single ovary in
females
- Ovary
- most birds have only left ovary but 2 ovaries
are typical of many raptors
- contains from 500 to several thousand primary
oocytes
- Testes & follicles increase dramatically in size as the
breeding season approaches.
- As day length increases, photic stimulation of
the hypothalamus results in the secretion of Gonadotropin releasing
hormone (GnRH below). When activated by GnRH, the anterior pituitary
secretes two gonadotropin hormones, follicle-stimulating hormone (FSH)
and luteinizing hormone (LH). FSH acts on sperm-producing structures in
the testes, while LH acts on the interstitial cells of the testes causing
them to secrete the steroid hormone testosterone. The pituitary gland
monitors the amount of testosterone in the blood, thus creating a negative
feedback loop to maintain hormone levels within a set range (Akins and
Burns 2001).
- Ambient visual cues, such as daylight, activate
photosensitive loci in the brain both indirectly, through the eyes, and
directly, through the skull. The hypothalamus of the bird brain contains
special cells that are sensitive to extremely low light levels,
intensities comparable to the amount of light that can penetrate brain
tissue (Akins and Burns 2001).

From: Akins and Burns (2001)
The pattern of testosterone secretion in free-living populations
of Song Sparrows.
Plasma levels peak in April and May as breeding got underway and then were
maintained at a lower “breeding baseline” during the
rest of the breeding season. As prebasic molt ensued, plasma levels of
testosterone were basal and remained so throughout autumn and winter.
From: Wingfield and Hahn (1994).
Biological
actions of the steroid hormone testosterone. The morphological, physiological and behavioral actions
of testosterone that are essential
for male reproductive function are given on the right hand and lower sides of
the figure. The “costs” of prolonged high levels of testosterone are given on
the
left hand side in italics. The patterns of plasma testosterone levels may be a
function of secretion patterns to maintain male reproductive function, and
“costs”
of testosterone that require that plasma levels be low. From Wingfield et al.
(2000).
Testosterone
increases availability of carotenoids --
Androgens and carotenoids play a fundamental role in the expression of
secondary sex traits in animals that communicate information on individual
quality. In birds, androgens regulate song, aggression, and a variety of
sexual ornaments and displays, whereas carotenoids are responsible for the
red, yellow, and orange colors of the integument. Parallel, but independent,
research lines suggest that the evolutionary stability of each signaling
system stems from tradeoffs with immune function: androgens can be
immunosuppressive, and carotenoids diverted to coloration prevent their use
as immunostimulants. Despite strong similarities in the patterns of sex, age
and seasonal variation, social function, and proximate control, there has
been little success at integrating potential links between the two signaling
systems. These parallel patterns led us to hypothesize that testosterone
increases the bioavailability of circulating carotenoids. To test this
hypothesis, Blas
et al. (2006) manipulated
testosterone levels of Red-legged Partridges ( Alectoris rufa) while
monitoring carotenoids, color, and immune function. Testosterone treatment
increased the concentration of carotenoids in plasma and liver by >20%.
Plasma carotenoids were in turn responsible for individual differences in
coloration and immune response. These results provide experimental evidence
for a link between testosterone levels and immunoenhancing carotenoids that ( i)
reconciles conflicting evidence for the immunosuppressive nature of androgens,
( ii) provides physiological grounds for a connection between two of
the main signaling systems in animals, ( iii) explains how these
signaling systems can be evolutionary stable and honest, and ( iv) may
explain the high prevalence of sexual dimorphism in carotenoid-based
coloration in animals.
|

Red-legged Partridge (Photo by G. Bortolotti)
|
Sperm production
- occurs in seminiferous tubules of the testes (shown below)
- occurs best at slightly cooler temperatures, so spermatogenesis
may occur primarily at night when body temperatures are slightly lower

Light photomicrograph of a
section of a testis showing a seminiferous tubule during full
semen production. SG
indicates spermatogonia; PS, primary spermatocyte; Ss, secondary spermatocyte;
MS, mature spermatocyte; and
L, lumen (original magnification ×800) (Samour 2002).
- sperm are stored at the terminal end of the vas
deferens (seminal glomus), and this creates a swelling called the cloacal
protuberance
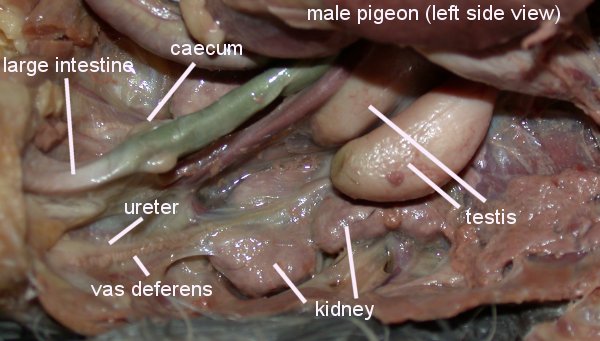
Male birds have paired abdominal testes lying
cranioventral to the first kidney lobe. Testes increase dramatically in size
during the breeding season. The vas deferens emerges medially and passes
caudally to the cloaca where it has a common opening with the ureter in the
Urodeum. The terminal vas deferens is swollen as a storage organ: the seminal
glomus (or seminal vesicle as in the drawing to the right).
As in mammals, sperm formation is temperature sensitive, and maturation is
assisted by nocturnal drops in temperature, or by the development of
scrotal-like external thermoregulatory swellings holding the seminal
glomera.
In addition, male birds tend to have relatively low extragonadal sperm
reserves and sperm are ejaculated soon after production in the testes. |
|

Cloacal protuberance
|
 Longitudinal section of the cloaca of a male
budgerigar during the
Longitudinal section of the cloaca of a male
budgerigar during the
culmination phase of the breeding cycle. SG indicates seminal glomus;
P, proctodeum; and C, cloaca (original magnification ×12) (Samour 2002).
|
Sperm
competition and testes size --
Comparative analyses suggest that a variety of ecological and behavioural
factors contribute to the
tremendous variability in extrapair mating among birds. In an analysis of 1010
species of birds, Pitcher et al. (2005) examined several ecological
and behavioural factors in relation to testes size; an index of sperm
competition and the extent of extrapair mating. In univariate and multivariate
analyses, testes size was significantly larger in species that breed colonially
than in species that breed solitarily, suggesting that higher breeding
density is associated with greater sperm competition. After controlling for
phylogenetic effects and other ecological variables, testes size was also
larger in taxa that did not participate in feeding their offspring. In analyses
of both the raw species data and phylogenetically independent contrasts,
monogamous taxa had smaller testes than taxa with multiple social mates, and
testes size tended to increase with clutch size, which suggests that
sperm depletion may play a role in the evolution of testes size. These results
suggest that traditional ecological and behavioural variables, such as
social mating system, breeding density and male parental care can account for a
significant portion of the variation in sperm competition in birds.
Testis size increases with colony size in Cliff
Swallows -- By using a
sample of over 800 male Cliff Swallows (Petrochelidon pyrrhonota) that
died during a rare climatic event in their Nebraska study area in 1996, Brown
and Brown (2003) investigated how testis size was related to body size, age,
parasite load, and a bird's past colony-size history. Testis volume increased
with body size. After correcting for body size, testis volume was lowest for
birds age 1 and 2 years but did not vary with age for males 3 years old or
more. Birds occupying parasite-free (fumigated) colonies had significantly
larger testes than did birds at nonfumigated sites. Testis volume increased
significantly with the size of the breeding colonies a bird had used in the
past. These results show within a species that larger testes are favored in
more social environments, probably reflecting a response to increased rates
of extrapair copulation (and thus sperm competition) among Cliff Swallows in
large colonies. The presence of ectoparasites, by inflating levels of plasma
corticosterone, may in turn reduce testis mass. These data provide no support
for the hypothesis that large testes, perhaps by producing more testosterone,
are immunosuppressive and thus costly for that reason.
|
|
Cliff Swallows
Temperature and the
timing of reproduction -- Many bird species reproduce earlier in
years with high spring temperatures, but little is known about the causal
effect of temperature. Temperature may have a direct effect on timing of
reproduction, but the correlation may also be indirect, for instance via food
phenology. As climate change has led to substantial shifts in timing, it is
essential to understand this causal relationship to predict future impacts of
climate change. Visser
et al. (2009)tested the direct effect of temperature on laying dates in
Great Tits (Parus major) using climatized aviaries in a 6-year
experiment. Temperature patterns from two specific years in which the wild
population laid either early (‘warm’ treatment) or late (‘cold’ treatment) were
mimicked. Laying dates were affected by temperature directly. Because the
relevant temperature period started three weeks prior to the mean laying date,
with a range of just 4°C between the warm and the cold treatments, and because
the birds were fed ad libitum, it is likely that temperature acted as a cue
rather than lifting an energetic constraint on the onset of egg production.
Visser et al. (2009) also found a high correlation between the laying dates of
individuals reproducing both in aviaries and in the wild, validating
investigations of reproduction of wild birds in captivity. These results
demonstrate that temperature has a direct effect on timing of breeding, an
important step towards assessing the implication of climate change on seasonal
timing.
Egg production
- Most birds have only one ovary and one oviduct. In early stages of
embryonic development, each female bird has two ovaries; only the left one
develops into a functional organ. In some birds, such as hawks, the right
ovary and oviduct usually develop. A mature ovary looks like a cluster of
grapes. and may contain up to 4,000 small ova which can develop into
mature ova.
- With fertilization, the ovum
(egg) becomes a
developing embryo
- The
embryo passes through the oviduct; typically takes about 24 hours (for
passerines & most other birds)
- The demand for calcium to make the egg shell is very high, and so
the circulating levels of blood calcium in birds are greatly elevated
compared to mammals, typically twice as much.
In most birds, only the left ovary
and oviduct persist. The ovary enlarges greatly during the breeding season.
Active ovaries resemble bunches of tiny grapes -- the developing follicles.
The oviduct opens medially to it in a funnel-shaped ostium. Ovulation results
in the release of an egg from a mature follicle on the surface of the ovary.
The egg, with extensive food reserves in the form of concentric layers of yolk,
is picked
up by the ostium and ciliary currents carry
it into the magnumregion.
Over about three hours the egg receives a coating of albumen.
The egg then passes into the isthmus,
where the shell membranes are deposited. This takes about one hour. The egg
them moves to the uterus, or shell gland, where
the calcareous shell is added and, in some birds, pigment is added in
characteristic patterns. The egg then passes into the vagina and cloaca for
laying.
|
|

Variation among bird species in the relative amount of yolk in eggs and the
amount of energy available to the developing embryo (kJ-g -1, or kilojoules per
gram). From top to bottom, the hatchlings are an altricial Brown Creeper, a
semiprecocial Least Tern, a precocial Ruddy Duck, a superprecocial Mallee Fowl
(Leipoa ocellata), and a Brown Kiwi (Apteryx australis).
Kiwis are ‘outliers.’ Female kiwis produce extremely large eggs for their size
(with substantial amounts of yolk), but young typically remain in the nest for
several days and so are best classified as semiprecocial (From: Sotherland and
Rahn 1987).
Kiwi lays an egg
|
The left egg found inside the female
oviraptorosaurian. The
texture of the shell pieces probably resembles the original
texture of the egg. Credit: Yen-nien Cheng
|
Eggs discovered inside dinosaur -- The discovery of eggs inside a
dinosaur has provided new clues about dinosaur reproductive biology and more
support for the hypothesis that birds evolved from dinosaurs. The pair of
eggs are the first found inside a dinosaur. Sato et al. (2005) found that the
dinosaur produced eggs in some ways like a crocodile and in other ways like a
bird. Crocodiles have two ovaries enabling them to lay a clutch of eggs.
Birds have a single ovary and lay only one egg at a time. The dinosaur's
egg-producing capability lay somewhere in between, suggesting a possible link
with modern birds. It had two ovaries, but produce only one egg at a time
from each ovary.
Sato et al. (2005) studied a dinosaur from a group of
dinosaurs called oviraptorosaurians. This type of dinosaur — probably 3 - 4
meters long — is a subgroup of the theropods. The dinosaur was excavated in China. The
similar size of the eggs suggests the creature's two oviducts each produced a
single egg at the same time.
|
Dinosaur egg
Female
birds can bias the sex of their chicks.-- Whether a bird is more
likely to lay a male or female egg depends on which sex will have the
greatest chance of doing well. Rutstein et al. (2004) adjusted the food
intake of female Zebra Finches [see photo of female (left) and male (right) Zebra
Finches below right] & found that well-fed females were more likely to
produce daughters, while less well nourished birds were more likely to have
sons. This is exactly as predicted by the fact that female offspring need to
be better nourished than males if they are to survive and grow well. 
The authors noted that: “In most animals sex
ratio is close to 50:50 and extremely resistant to change. In mammals,
including humans, the sex of the baby is determined by whether the sex
chromosome in the sperm is male or female. But in birds, it is the female’s
egg rather than the male’s sperm that determines what sex the chick will be.
Thus the female has the potential to determine the sex of her young by
whether she ovulates male or female eggs. In some way, female Zebra Finches
seem to be able to exert control over whether to produce a male or female egg
depending on which of the two is most likely to be successful. Our research
tells us that they do it, and we understand why. The big question is: how do
they do it?”
In many animals, females need to be
well-nourished and in good condition if they are to breed, as eggs are costly
to produce. Bigger eggs tend to lead to bigger young that are more likely to
survive. Such ‘sex ratio adjustment’ is well documented in certain insects,
such as bees and wasps, but is less well understood in birds and
mammals.
Birds are an excellent model to use in the
study of sex ratio adjustment because, using molecular techniques, scientists
can establish the sex of each egg soon after laying. Further, all the
resources given to the developing embryo are present in the egg at laying.
Thus the size and the content of the egg are measures of the amount of
resources that the female has allocated to that egg, which affects its
subsequent survival chances.
The authors explained: “We manipulated the diet
quality of Zebra Finches to look at the effects of body condition on female
investment. We found that females were able to exert a strong degree of
control over the production of male and female eggs. When females were fed on
a low quality diet, they laid eggs that were considerably lighter than those
laid when they were fed on a high quality diet, and they also laid far more
male eggs on a low quality diet. This is the converse situation to that
described 30 years ago for mammals, but it makes sense for Zebra Finches.
Previous research has shown that under poor nutritional conditions, female
Zebra Finches grow more slowly and survive less well compared to males. Therefore,
females are producing more of the sex with the highest survival chances under
those conditions.”
|

Two potential mechanisms for determination among birds. (A) the
presence of the W chromosome triggers femaleness or (B) the presence of two Z
chromosomes confers maleness.
|
Avian sex determination (Ellegren 2001) -- The molecular
determinants behind sexual development in birds remain a mystery. The process
is known to be different from that in mammals, with no homolog to the gene
that confers maleness in mammals found in birds. The failure to identify such
a gene in birds is probably a reflection of the fact that, despite the
occurrence of two sexes being nearly universal throughout the animal kingdom,
the genes involved seem virtually unrelated among metazoan phyla. These
differences raise obstacles for comparative or candidate gene approaches in
studies of sexual development.
In birds, females are the heterogametic sex,
with one copy each of the Z and W sex chromosomes. Males are homogametic
(ZZ). However, it is not clear whether it is the presence of the
female-specific W chromosome that triggers female development, or the dose of
Z chromosome that confers maleness. An intriguing additional
possibility is that both Z and W matter! In marsupials, for example, Y acts
as a dominant testis determining chromosome, while the X chromosome
determines the choice between pouch and scrotum. Maybe a system where the two
sex chromosomes mediate different aspects of sex differentiation is also used
in birds.
|

Vertebrate sex determination systems. Phylogeny of major vertebrate clades
showing the sex determining systems
found in members of the respective clade. ‘Multiple’ indicates involvement of
more than one pair of chromosomes in sex determination.
TSD: temperature-dependent sex determination (From: Ezaz 2006).
Incubation temperature
and avian sex ratios -- Although common in reptiles, incubation
temperature has not been considered to be a factor in determining sex ratios  in birds. However, Goth and Booth (2005)
found that incubation temperature does affect sex ratios in megapodes, which
are exceptional among birds because they use environmental heat sources for
incubation. In the Australian
Brush-turkey (Alectura
lathami), a mound-building megapode, more males hatch at low incubation
temperatures and more females hatch at high temperatures, whereas the
proportion is 1:1 at the average temperature found in natural mounds. Chicks
from lower temperatures weigh less, which probably affects offspring survival,
but are not smaller. Megapodes possess heteromorphic sex chromosomes like other
birds, which eliminates temperature-dependent sex determination, as described for
reptiles, as the mechanism behind the skewed sex ratios at high and low
temperatures. Instead, Goth and Booth (2005) suggest a sex -biased
temperature-sensitive embryo mortality because mortality was greater at the
lower and higher temperatures, and minimal at the middle temperature where the
sex ratio was 1:1.
in birds. However, Goth and Booth (2005)
found that incubation temperature does affect sex ratios in megapodes, which
are exceptional among birds because they use environmental heat sources for
incubation. In the Australian
Brush-turkey (Alectura
lathami), a mound-building megapode, more males hatch at low incubation
temperatures and more females hatch at high temperatures, whereas the
proportion is 1:1 at the average temperature found in natural mounds. Chicks
from lower temperatures weigh less, which probably affects offspring survival,
but are not smaller. Megapodes possess heteromorphic sex chromosomes like other
birds, which eliminates temperature-dependent sex determination, as described for
reptiles, as the mechanism behind the skewed sex ratios at high and low
temperatures. Instead, Goth and Booth (2005) suggest a sex -biased
temperature-sensitive embryo mortality because mortality was greater at the
lower and higher temperatures, and minimal at the middle temperature where the
sex ratio was 1:1.
Copulation &
fertilization:
- For most birds, copulation involves a 'cloacal kiss', with the
male on the female's back & twisting his tail under the female's
- copulation typically lasts just a few seconds (but there are
exceptions - see Phony phallus puts sperm ahead in bird first below)
Bald Eagles mating
White-throated Kingfishers mating
Phony
phallus puts sperm ahead in bird first-- "These birds
would be at it for 10-20 minutes," said co-author Tim Birkhead of the
Red-billed Buffalo Weaver. Males use their organ to rub females and improve
their sperm's chance of success. Few male birds have a phallus; most achieve
fertilization via a cloacal kiss. So 19th-century reports of a mock member in
the Buffalo Weaver sent Winterbotton et al. (2001) to Namibia. Catching
the birds in the act was tough, recounts Birkhead: "In 3 years we saw
eight matings." Pairs occasionally emerged from nests and flew to a
nearby tree. "I'd run after them, sweating profusely with my binoculars
steaming up," he says. The pair would start bouncing up and down - over
numerous consecutive bouts. Compared to the 1-2 second tryst of most birds,
their staying power is unique. Yet, entry of the elusive organ was hard to
make out. Even in captivity "they performed beautifully," but the
view was blocked, says Birkhead. So they glued a piece of cardboard to an
unlucky bird's member. This did not prevent mating, suggesting that the
Buffalo Weaver organ is actually a weapon in sperm wars. By choosing a male
who rubs longest or best, females may be selecting top-quality sperm.
Paternity testing revealed that female Buffalo Weavers sire birds from
multiple males, providing evidence of sperm competition. Time spent courting
must be shown to predict sperm transfer or success to really back up the
idea. The 1.5-cm appendage lacks blood vessels and has a twisted furrow down
its length. Males in communal nests have longer ones than those that live
alone, showing that size is a factor in social success. But for males at
least, the phallus is for more than foreplay. -- Helen Pearson, Nature
Science Update
|
|
- males in a few species, including most waterfowl & ostriches
(see diagram below), have an intromittent organ; most males do not

Diagram of the left lateral view of a retracted and erect phallus of a male Emu
or Rhea. The top drawing
represents the phallus within the pouch. A. vas
deferens, B. urideum, C. proctodeum, D. pocket to contain phallus,
E. erectile wall of phallus, F inverted hollow tube of phallus, G. phallic
sulcus, H. erectile tissue, and I. erect phallus
with blind hollow tube. (Source: http://www.cassowary.com/workshop.html)

Examples of genital covariation in waterfowl.
(A) Harlequin Duck (Histrionicus histrionicus) and (B) African Goose (Anser
cygnoides), two species with a short phallus and no forced copulations, in
which females have simple vaginas. (C) Long-tailed Duck (Clangula hyemalis),
and (D) Mallard (Anas
platyrhynchos), two species
with a long phallus and high levels of forced copulations, in which females
have very elaborate vaginas (size bars = 2 cm). ] = Phallus, * = Testis, star =
Muscular base of the male phallus, > = upper and lower limits of the vagina
(From: Brennan
et al. 2007).
Eversion of a male Muscovy duck penis
Explosive
eversion and functional morphology of the duck penis -- Coevolution of male and female genitalia
in waterfowl has been hypothesized to occur through sexual conflict. This
hypothesis raises questions about the functional morphology of the waterfowl
penis and the mechanics of copulation in waterfowl. Brennan et al. (2010) used
high-speed video of phallus eversion and histology to describe for the first
time the functional morphology of the avian penis. Eversion of the 20 cm
muscovy duck penis is explosive, taking an average of 0.36 sec, and achieving a
maximum velocity of 1.6 m sec−1. The collagen matrix of the penis is
very thin and not arranged in an axial-orthogonal array, resulting in a penis
that is flexible when erect. To test the hypothesis that female genital
novelties make intromission difficult during forced copulations, Brennan et al.
(2010) investigated penile eversion into glass tubes that presented different
mechanical challenges to eversion. Eversion occurred successfully in a straight
tube and a counterclockwise spiral tube that matched the chirality of the
waterfowl penis, but eversion was significantly less successful into glass
tubes with a clockwise spiral or a 135° bend, which mimicked female vaginal
geometry. These results support the hypothesis that duck vaginal complexity
functions to exclude the penis during forced copulations, and coevolved with
the waterfowl penis via antagonistic sexual conflict.
Near the junction of the vagina
and shell gland of female birds are deep glands lined with simple columnar
epithelium. These are the sperm storage tubules, so called because they can
store sperm for long periods of time (10 days to 2 weeks). After an egg is
laid, some of these sperm may move out of the tubules into the lumen of the
tract, then migrate farther up to fertilize another egg.
 Avian sperm
Avian sperm
|
 Avian sperm storage tubules
Avian sperm storage tubules
|

Photomicrographs of sperm storage tubules contained stained and unstained
spermatozoa from domestic chicken (Gallus
domesticus)
hens (a, b) and turkey (Meleagris gallopavo) hens (c, d). Arrows
indicate stained spermatozoa; arrowheads designate unstained spermatozoa.
Scale bars = 25 micrometers. From: King et al. (2002).
King et al. (2002) found that spermatozoa from two different
inseminations (one with stained sperm, one with unstained sperm) generally
segregated into different storage tubules in both chicken and turkey hens.
Storage tubules contained mixed populations of spermatozoa were
found in only 4% of chicken and 12% of turkey storage tubules examined. They
concluded that the mechanism of last-male precedence does
not appear to be due to the stratification of spermatozoa within the tubules.
|
|
Innervation of sperm storage tubules (Freedman et al. 2001) --
Immunohistochemical staining of a turkey uterovaginal junction and sperm
storage tubules. This micrograph shows a fluorescing neuron (green) near some
sperm storage tubules (SST). The blue areas (se) are the surface epithelium
lining the lumen of the uterovaginal junction and the epithelium of the sperm
storage tubules. The arrow points to a magenta-stained area of one SST that
indicates the presence of actin (a protein found in smooth muscle. The total
image is 19 micrometers across.
This association between neurons and SSTs provides evidence that SSTs are
innervated and suggests that the storage and release of sperm from SSTs can,
perhaps, be controlled. |
Post-insemination
events (Birkhead and Brillard 2007) -- Most birds do not have a phallus
and, in these species, insemination occurs via the so-called ‘cloacal kiss.’
Depending on taxa, sperm are ejaculated into the cloaca or vagina and rely on
their motility to reach the numerous sperm-storage tubules (SSTs) located at
the junction of the vagina and the uterus. As a consequence of selection during
their migration through the vagina, only 1–2% of inseminated sperm enter the
SSTs, the rest are probably ejected the next time that the female defecates.
The SSTs contain only morphologically normal sperm, suggesting either that only
normal sperm successfully traverse the vagina or that only normal sperm are
‘accepted’ by the SSTs. An unknown but probably small proportion of sperm move
directly to the infundibulum (the site of fertilization) without entering the
SSTs, although these are likely to fertilize only a single ovum.
That sperm in the SSTs are invariably positioned
with their heads directed towards the distal end of the tubule suggests that
egress from the SSTs is passive. Sperm are lost from the SSTs more or less
continuously at a constant per capita rate. They enter the uterus and are
carried passively to the infundibulum. Sperm accumulate or move relatively
slowly through the infundibulum so that there is usually a population available
to fertilize each ovum as it is ovulated. On ovulation, the ovum is captured by
the prehensile, funnel-shaped infundibulum and the sperm swarm over the surface
of the ovum; their target is the germinal disc, which contains the female
pronucleus. At this stage, the ovum is bounded by the inner perivitelline
layers (IPVL). The clustering of sperm and holes made by sperm in the IPVL
around the germinal disc suggest that sperm might use chemical signals to
locate the germinal disc.
In contrast to most other taxa, where only a
single sperm enters the ovum, polyspermy is typical in birds. Several sperm
enter the germinal disc region, hydrolyzing the IPVL via the acrosome reaction
of the sperm, whereby the release of enzymes from the sperm acrosome enables
the sperm nucleus to enter the ovum. However, only a single spermatozoon fuses
with the female pronucleus and the remaining sperm are shifted to the periphery
of the germinal disc and play no further part in development. Fertilization
includes the penetration of ovum by sperm as well as the fusion of the male and
female pronuclei (syngamy). Because embryo development begins almost
immediately, many cell divisions have occurred by the time the ovum has become
incorporated into the egg and the egg is laid (in most species) 24 hr later.

Scanning electron photomicrograph of a
budgerigar spermatozoon. A indicates
acrosome; H, head; and T, tail (original
magnification ×20000) (Samour 2002).
|

Transmission electron photomicrograph of the longitudinal
section of part of the nucleus and midpiece of a Budgerigar
spermatozoon. N indicates nucleus; PC, proximal centriole;
DC, distal centriole; F, axial filament complex; and
M, mitochondria (original magnification ×30000) (Samour 2002).
|
Fertilization of the egg usually
occurs in the infundibulum.

Light photomicrograph of a zona-free hamster ovum with
numerous budgerigar spermatozoa bound to its surface
(original magnification ×215) (Samour 2002).
Repelling clingy exes helps snipe save sperm -- Writer Gore Vidal once said that
he never passed up an opportunity to have sex or appear on television. Some
male birds would disagree on at least one count. Having mated with a female,
a male Great Snipe ( Gallinago media) will reject her further advances
and even chase her away. Male Great Snipe form leks to eye up the talent
before choosing a mate. A few males get the most sex. Popular birds can get
more than half of the matings, perhaps 10 a day. Hence their pickiness,
suggest Saether et al. (2001). As male Great Snipe take no part in caring for
their offspring, it was thought they had nothing to lose by mating as much as
possible. But top males, overburdened with potential partners, must share
sperm with care and spread their favors around. Sperm budgeting is the only
possible explanation for male snipes' ungrateful behavior. Like a
nightclub, Great Snipe leks see their share of aggravation. "All four
kinds of mating conflicts happen" - male choice, female choice, and male
and female competition - explains Saether. Males are more likely to repel
clingy exes if there are a lot of other females around. Females fight with
one another, and males from neighboring territories chase their rivals'
females away. Hostility towards old flames might be a bid to maintain order.
"If a male gets rid of an unwanted female it's one less problem to worry
about," says Saether. Female snipe probably seek to mate again so that
they can get enough sperm to fertilize their eggs. Rejected females tend to
lower their sights and settle for less popular males. -- John Whitfield, Nature
Science Update
|
|
The Avian
Egg
Birds' eggs, like the birds
themselves, vary enormously in size. The largest egg from a living bird belongs
to the ostrich. It is over 2000 times larger than the smallest egg
produced by a hummingbird (see photo to the right; Source:http://www.pma.edmonton.ab.ca/vexhibit/eggs/vexhome/sizeshap.htm). Ostrich eggs are about 180 mm long and
140 mm wide and weigh 1.2 kg. Hummingbird eggs are 13 mm long and 8 mm wide and
they weigh only half of a gram. The extinct Elephant Bird from Madagascar
produced an egg 7 times larger than that of the Ostrich! Within the egg, three
extraembryonic membranes support the life & growth of the embryo:
2000 times larger than the smallest egg
produced by a hummingbird (see photo to the right; Source:http://www.pma.edmonton.ab.ca/vexhibit/eggs/vexhome/sizeshap.htm). Ostrich eggs are about 180 mm long and
140 mm wide and weigh 1.2 kg. Hummingbird eggs are 13 mm long and 8 mm wide and
they weigh only half of a gram. The extinct Elephant Bird from Madagascar
produced an egg 7 times larger than that of the Ostrich! Within the egg, three
extraembryonic membranes support the life & growth of the embryo:
- amnion
- surrounds only the embryo
- inner layer of cells secretes amniotic fluid in
which the embryo floats; fluid keeps the embryo from drying out and
protects it
- chorion - surrounds all embryonic structures & serves as a
protective membrane
- allantois (or allantoic sac)
- grows larger as embryo grows, fuses with the
chorion & is called the chorio-allantoic membrane
- works together with chorion to permit
respiration (exchange of oxygen and carbon dioxide) and excretion
- important in storage of nitrogenous wastes (uric
acid)



Relative egg mass
(corrected for adult mass) is greater in species with longer embryonic periods
(days) among 64 passerine species in tropical Venezuela,
subtropical Argentina, and
north temperate Arizona.
Open symbols reflect cavity-nesting species and show an interacting effect
where their larger clutches are associated with relatively smaller eggs.
Egg size
variation among tropical and temperate songbirds -- Species with “slow” life history
strategies (long life, low fecundity) are thought to produce high-quality
offspring by investing in larger, but fewer, young. Larger eggs
are indeed associated with fewer eggs across taxa and can yield
higher-quality offspring. Tropical passerines appear to follow theory because
they commonly exhibit slow life history strategies and produce larger, but
fewer, eggs compared with northern species. Martin
(2008) found that relative egg mass
(corrected for adult mass) varies extensively in the tropics and subtropics for
the same clutch size, and proposed a hypothesis to explain egg size variation both within the tropics
and between latitudes: Relative egg mass
increases in species with cooler egg temperatures
and longer embryonic periods to offset associated increases in energetic
requirements of embryos. Egg temperatures
of birds are determined by parental incubation behavior and are often cooler
among tropical passerines because of reduced parental attentiveness of eggs.
Cooler egg temperatures
and longer embryonic periods explained the enigmatic variation in egg mass within and among regions, based
on field studies in tropical Venezuela
(36 species), subtropical Argentina
(16 species), and north temperate Arizona
(20 species). Alternative explanations were not supported. Thus, large egg sizes may reflect compensation for
increased energetic requirements of cool egg temperatures
and long embryonic periods that result from reduced parental attentiveness in
tropical birds.
Egg composition and hatchling phenotype -- Parental investment in eggs and,
consequently, in offspring can profoundly influence the phenotype, survival
and evolutionary fitness of an organism. Avian eggs are excellent model
systems to examine maternal allocation of energy translated through egg size
variation. Dzialowski1and Sotherland (2004) used the natural range in Emu (Dromaius
novaehollandiae) egg size, from 400 g to >700 g, to examine the
influence of maternal investment in eggs on the morphology and physiology of
hatchlings. Female Emus provisioned larger eggs with a greater absolute
amount of energy, nutrients and water in the yolk and albumen. Variation in
maternal investment was reflected in differences in hatchling size, which
increased isometrically with egg size. Egg size also influenced the
physiology of developing Emu embryos, such that late-term embryonic metabolic
rate was positively correlated with egg size and embryos developing in larger
eggs consumed more yolk during development. Large eggs produced hatchlings
that were both heavier (yolk-free wet and dry mass) and structurally larger
(tibiotarsus and culmen lengths) than hatchlings emerging from smaller eggs.
As with many other precocial birds, larger hatchlings also contained more
water, which was reflected in a greater blood volume. Emu maternal investment
in offspring, measured by egg size and composition, is significantly
correlated with the morphology and physiology of hatchlings and, in turn, may
influence the success of these organisms during the first days of the
juvenile stage.
|
|
- Eggs consists of 4 primary components:
- yolk
- energy-rich supply of food
- 21 - 36% lipids & 16 - 22% proteins (with
the rest being water)
- the yolk is suspended in the center of the egg
by twisted strands of protein fibers called chalazae (shown below)

Yolk contains maternal antibodies -- Antibodies are deposited in eggs
during yolk formation through the deposition of immunoglobulins, primarily
IgY (also called IgG), in the yolk. In Chickens (Gallus domesticus),
maternal IgY is catabolized by offspring over the first 14 days post-hatching
and, by about 5 days post-hatching, offspring begin to synthesize their own
IgY. As a result, after approximately two weeks the circulating IgY in young
is principally of endogenous origin. Adult levels are attained between six
weeks and six months of age. However, maternal antibodies may continue to
affect offspring phenotype even after they are catabolized by influencing
growth and developmental rates. In the absence of maternal IgY in chickens
(due to surgical bursectomy of the mother during her own embryogenesis), the
number of cells in the spleen that help lymphocytes (helper T cells) attack
antigens (foreign proteins on pathogens) is depressed. Also, the immune
responsiveness of offspring is depressed, which could lower the survival of
offspring particularly in harsh disease environments (Grindstaff et al.
2003).
|

Antibodies 'attack' pathogens or toxins they produce by binding to antigens (e.g.,
proteins in the membranes of
bacteria) via their 'binding sites' (the black areas above). This
binding can neutralize toxins and attract white blood cells that eliminate pathogens (by
phagocytosis).
|

Maternal secretion of antibodies and absorption by the young occur only
prenatally in birds (with the exception of pigeon crop milk)
(From: Boulinier and Staszewski 2008).
|
The familiar color of a chicken’s
egg yolk (a) is in stark contrast to the richly pigmented egg yolk of a
lesser Black-backed Gull, Larus
fuscus (b). Such high
maternal investment of carotenoids into egg yolk is typical among wild bird
species, suggesting that these biologically active pigments serve important
functions in the developing bird (From: Blount et al. 2000).
|
Why egg yolk is yellow (or red) (Blount et al. 2000) -- Egg yolk
in birds is colored yellowish-red by carotenoids. Until recently, there has
been no adaptive explanation of why many egg-laying animals provision their
eggs so richly with carotenoids. It now appears that, in developing birds,
carotenoids protect vulnerable tissues against damage caused by free
radicals. Athough embryonic tissues depend on oxidizable, unsaturated fatty
acids in yolk, their abundance makes the tissues susceptible to peroxidation
caused by reactive oxidative metabolites and by free radicals, which are
produced as normal by-products of metabolism. Protection against lipid
peroxidation in young birds is afforded by the actions of yolk-derived
carotenoids and other antioxidants, like vitamin E. Antioxidants also protect
passively-acquired antibodies (IgY; see above) against break-down. Thus,
maternal investment in egg composition, including carotenoids, might have a
greater influence on offspring viability than has been realized. The use of
carotenoid pigments in the sexual displays of female birds might indicate
their ability to produce high quality eggs and chicks.
|
- albumen
- 90% water & 10% protein
- the embryo's water supply, but also serves as a
'shock-absorber' to help protect the embryo
- buffers embryo from sudden changes in
temperature
- shell membranes
- attached to the shell are two membranes, the
inner and outer shell membranes. They protect the egg from bacterial
invasion and help prevent rapid evaporation of moisture from the egg.

Keratin fibers from the outer
shell membrane can be seen above, attached to the
calcium carbonate crystals
that make up the main shell structure.
(Source: http://www.rit.edu/~tld0898/SEM.html)
- shell
- protects the embryo
- contains thousands of pores (see diagram below)
that permit gas exchange
- generally white in cavity-nesters & colored
and patterned in open nesters (see Ecology of egg colors below)
- color is added to the eggshell from pigments
secreted by cells in the wall of the uterus

Weaker
Birds Use Steroids to Boost Offspring -- Verboven et al. (2003) reported
that female gulls in poor condition were more likely to give their chicks a
hormone boost to improve their chances of survival. Verboven and her
colleagues experimentally enhanced maternal condition by supplementary
feeding Lesser
Black-backed Gulls ( Larus
fuscus) during egg formation and compared the concentrations of steroids
(including testosterone) in their eggs with those in eggs laid by control
females. Egg androgens could affect offspring performance directly through
chick development and/or indirectly through changes in the competitive
ability of a chick relative to its siblings. Contrary to expectation, females
with experimentally enhanced body condition laid eggs with lower levels of
androgens. This suggests that less healthy females pass on more steroids than
healthy ones in a bid to enhance the performance of their young. Verboven
noted that “We originally thought that gulls in good condition would
put more steroids in their eggs. But we discovered that healthy birds don’t
tend to give their eggs the extra boost.” She compared the situation to
struggling athletes who take performance-enhancing drugs. She added: “A poor
sports person maybe wants to use steroids to conceal poor performance but if
you are good you don’t need to use them.”
|
|
Avian mothers create different phenotypes by
hormone deposition in their eggs --
In birds, mothers deposit substantial amounts of androgens in their eggs, and
experimental evidence indicates that these maternal androgens influence the
chick's early development. Despite the well-known organizing role of sex
steroids on brain and behavior, studies on avian maternal egg hormones almost
exclusively focus on the chick phase. Eising et al. (2005) found that, in
Black-headed Gulls, maternal androgens in the egg enhance the development of
the nuptial plumage and the frequency of aggressive and sexual displays (see
Figure above) almost 1 year after hatching. The long-lasting effects may be
mediated by an upregulation of androgen receptors later in life.
Alternatively, the early hormone exposure may have influenced the hypothalamus-pituitary-gonad
axis, resulting in higher androgen production later in life. The long-lasting
effects of egg androgens are almost certainly beneficial for Darwinian
fitness. Successful territory establishment and defense by means of
aggressive interactions are essential for reproductive success in this
colonial breeder. In addition, the displays are important for mate selection.
Clearly, in birds, maternal hormone deposition in eggs may profoundly
influence individual differentiation of fitness related traits. Since these
hormones suppress early immune function of the chick and reduce long-term
survival, mothers may be faced with a trade-off between producing offspring
with lower survival prospects but higher reproductive success per year, or
with higher chances of survival and lower annual reproductive output. By
producing eggs that differ in levels of maternal hormones, mothers seem to
produce a variety of phenotypes, perhaps an adaptive strategy in
unpredictable environmental conditions. Since natural selection acts upon
such phenotypic variation, shaping a population's demography, the role of
maternal androgens in this selective process may be much greater than
anticipated until now.
|
The Ecology of Egg Colors (see Birds: A
Virtual Exhibition - The
Provincial Museum of Alberta)
Egg colors and markings have
strong adaptive values. Originally, birds' eggs were probably all white, as
reptile eggs are. Eggs that are laid on the ground or in open nests in trees,
rather than in cavities, often exhibit cryptic coloration. The eggs blend in
with their surroundings and are much less visible to potential predators (e.g.,
aKilldeer nest).
Sometimes eggs that are laid in
open nests are white at first. They then become stained by the mud and rotting
vegetation in the nest. Grebes lay white eggs that become stained and
cryptically colored over time.
In some species, such as the
Common Murre, where different females lay eggs with very different markings,
the uniqueness may have a purpose. Distinctive patterns, as in the eggs shown
below, help females identify their own egg in a colony where thousands of eggs
may dot a cliff face.
Eggs of kingfishers and other
cavity nesting birds, such as woodpeckers and some owls, are often white. The
brightness of the eggs may help the parents to more easily locate them in the
cavity. Shown here is the egg of a Barn Owl.
Evolution of egg color and patterning in
birds -- Avian
eggs differ so much in their color and patterning from species to species that
any attempt to account for this diversity might initially seem doomed to
failure. Kilner (2006) reviewed the literature that, when combined with the
results of some comparative analyses, suggests that just a few selective agents
can explain much of the variation in egg appearance. Ancestrally, bird eggs
were probably white and immaculate. Ancient diversification in nest location,
and hence in the clutch's vulnerability to attack by predators, can explain
basic differences between bird families in egg appearance. The ancestral white
egg has been retained by species whose nests are safe from attack by predators,
while those that have moved to a more vulnerable nest site are now more likely
to lay brown eggs, covered in speckles, just as Wallace hypothesized more than
a century ago. Even blue eggs might be cryptic in a subset of nests built in
vegetation. It is possible that some species have subsequently turned these
ancient adaptations to new functions, for example to signal female quality, to
protect eggs from damaging solar radiation, or to add structural strength to
shells when calcium is in short supply. The threat of predation, together with
the use of varying nest sites, appears to have increased the diversity of egg
coloring seen among species within families, and among clutches within species.
Brood parasites and their hosts have probably secondarily influenced the diversity
of egg appearance. Each drives the evolution of the other's egg color and
patterning, as hosts attempt to avoid exploitation by rejecting odd-looking
eggs from their nests, and parasites attempt to outwit their hosts by laying
eggs that will escape detection. This co-evolutionary arms race has increased
variation in egg appearance both within and between species, in parasites and
in hosts, sometimes resulting in the evolution of egg color polymorphisms. It
has also reduced variation in egg appearance within host clutches, although the
benefit thus gained by hosts is not clear.

Many small songbirds have eggs
with just a ‘ring’ of small spots around the broad end that does little to make
the eggs cryptic. Evidence now suggests that such spots are located where the
eggshell is a bit thinner (likely due to a calcium deficiency in the diet of
female birds), with the pigment serving to strengthen the shell (Gosler et al.
2005). The spots consist of protoporphyrin pigment that birds synthesize during
production of the heme component of hemoglobin (Burley and Vadhera 1989) and
integration of this pigment into the eggshell provides additional strength.
When a female bird has insufficient calcium to deposit in a shell,
protoporphyrin molecules that have a semi-crystalline structure similar to that
of eggshells are apparently deposited instead instead of calcium. As a result,
the spots occur precisely where the shell is a bit thinner.

Several species of birds have
blue eggs, and David Lack (1958) suggested that, in habitats where light levels
are low, blue eggs might be cryptic. If true, that could help explain the blue
eggs of some open-cup nesting birds that occur in forest habitats such as Wood
Thrushes. However, Lack’s hypothesis cannot explain why some birds that nest in
cavities, like European Starlings and Eastern Bluebirds, also have blue eggs.
One hypothesis is that the blue-green color of eggshells represents a signal of
female quality to their mates ( Moreno
and Osorno 2003). The pigment responsible for the blue-green color is
biliverdin, a substance produced when the hemoglobin of damaged red blood cells
is catabolized and also known to have strong antioxidant properties.
Antioxidants are important because they can convert free radicals, molecules
that can damage DNA, proteins, and other macromolecules, into less reactive
substances. Deposition of this pigment in eggshells by laying females may,
therefore, signal their capacity to produce antioxidants and control free
radicals. Male birds paired to females of such quality that they are able to
deposit antioxidants in eggshells rather than retaining them may then expend
greater effort in caring for their superior offspring (Kilner 2006). In support
of this hypothesis, the provisioning rates of male Pied Flycatchers (Ficedula
hypoleuca) and the intensity of the blue coloration of eggs were found to
be positively correlated (Moreno et al. 2004). Also, female Eastern Bluebirds
in better body condition were found to lay more colorful eggs, supporting the
hypothesis that biliverdin pigmentation in eggshells reflects female condition
(Siefferman et al. 2006).
Egg-laying:
"Initially the female stood
motionless in the nest cup. The first sign of approaching egg- laying was
usually intensified breathing, occasionally with rhythmic opening and closing
of the bill that pointed either horizontally forwards or more or less upwards.
The head was drawn in and the body feathers were somewhat fluffed out; the Coal
Tit in addition raised its crown feathers. The tail was kept horizontal or
elevated up to about 45 degrees”. Then the tip of the tail started nodding
movements synchronously with rhythmic depressions of the rump.These movements
which apparently were caused by throes of parturition when the egg traveled
down the oviduct, were almost invisible to begin with but gathered in strength
and ended with a sudden elevation of the rump that marked the moment of
egg-laying. Then the bird “froze” in a motionless posture, termed “recovery
phase.” This last rise of the rump clearly indicated that the egg had just been
laid.
Duration of egg-laying varies a
great deal even within species. The opening and closing of the bill and
rhythmic movements of the back and tip of the tail occurs repeatedly for up to
4 minutes in the Prairie Warbler, presumably corresponding to the duration of
egg- laying. For 3 eggs of the Goldcrest, only 8-9 seconds elapsed between the
first visible sign of pressure and the moment of egg- laying. In tits, this
period varied from about 10 to 77 seconds, mostlv 20-30 seconds. The Cuckoo (Cuculus
canorth) which is a brood parasite, is known to lay the egg remarkably
swift, usually within 10 seconds with a lower limit of only 3-4 seconds.
Presumably this short duration is an adaptation to its parasitic
behavior."--- From: Haftorn (1996).


Female birds turns part of the cloaca and the last segment of the
oviduct inside out ("like a glove"). The vent is then everted and the
egg emerges far outside at the end of the bulge. As a result, the egg does not
contact the walls of the cloaca and get contaminated by feces. In addition, the
intestine and inner part of the cloaca are kept shut by the emerging egg, and
their contents cannot leave when the hen strains to deliver the egg. Therefore,
eggs are always clean when laid (van der Molen 2002).
Wandering Albatross laying an egg
Kiwi laying an egg
Literature Cited:
Akins, C. & M. Burns. 2001.
Visual control of sexual behavior. In R. G. Cook (Ed.), Avian visual
cognition [On-line]. Available:www.pigeon.psy.tufts.edu/avc/akins/
Blas, J., L. Perez-Rodriguez, G.
R. Bortolotti, J. Vinuela, and T. A. Marchant. 2006. Testosterone increases
bioavailability of carotenoids: insights into the honesty of sexual signaling. Proceedings National Academy
of Sciences 103: 18633-18637.
Blount, J. D., D. C. Houston, and
A. P. Møller. 2000. Why
egg yolk is yellow. Trends in Ecology and Evolution 15: 47-49.
Boulinier, H. and V. Staszewski.
2008. Maternal
transfer of antibodies: raising immuno-ecology issues. Trends in Ecology
and Evolution, in press.
Brennan, P. L. R., C. J. Clark,
and R. O. Prum. 2010. Explosive
eversion and functional morphology of the duck penis supports sexual conflict
in waterfowl genitalia. Proceedings of the Royal Society B, online early.
Brennan, P. L., R. O. Prum, K. G.
McCracken, M. D. Sorenson, R. E. Wilson, and T. R. Birkhead. 2007. Coevolution
of male and female genital morphology in waterfowl. PLoS ONE 2: e418.
Brown, C.R. and M. B. Brown.
2003. Testis size increases with colony size in cliff swallows. Behavioral
Ecology 14:569-575.
Burley, R. W. and D. V. Vadhera.
1989. The avian egg. John Wiley, New
York, NY.
Dzialowskil, E. M. and P. R.
Sotherland. 2004. Maternal effects of
egg size on emu Dromaius
novaehollandiae egg
composition and hatchling phenotype. J. Exp. Biol. 207:597-606.
Eising, C. M., W.
Müller & T. G.G. Groothuis. 2005. Avian
mothers create different phenotypes by hormone deposition in their eggs.
Proc. Royal Soc. London, Biol. Letters: early on-line.
Ellegren, H. 2001. Hens, cocks
and avian sex determination: a quest for genes on Z or W? European Molecular Biology
Organization Reports 2:192-196.
Ezaz, T., R. Stiglec, F. Veyrunes,
and J. A. Marshall Graves. 2006. Relationships
between Vertebrate ZW and XY Sex Chromosome Systems. Current Biology 16:
R736-R743.
Freedman, S. L., V. G. Akuffo,
and M. R. Bakst. 2001. Evidence for the innervation of sperm storage tubules in
the oviduct of the turkey (Meleagris gallopavo). Reproduction 121:
809-814.
Gosler, A.G., J. P. Higham, and
S. J. Reynolds. 2005. Why are birds’ eggs speckled? Ecology Letters 8:
1105-1113.
Goth, A. and D. T. Booth. 2005. Temperature-dependent
sex ratio in a bird. Biology Letters 1: 31-33.
Grindstaff, J. L., E. D. Brodie
III, and E. D. Ketterson. 2003. Immune function across generations: integrating
mechanism and evolutionary process in maternal antibody transmission. Proc.
Royal Soc. Lond. B 270: 2309-2319.
Kilner, R. M. 2006. The
evolution of egg colour and patterning in birds. Biological Reviews 81:
383–406.
Haftorn, S. 1996. Egg-laying
behavior in tits. Condor 98:863-865.
King, L.M., J. P. Brillard, W.M.
Garrett, M.R. Bakst, and A.M. Donoghue. 2002. Segregation of spermatozoa within
sperm storage tubules of fowl and turkey hens. Reproduction 123:79-86.
Lack, D. 1958. The significance
of the colour of turdine eggs. Ibis 100: 145-166.
Martin, T. E. 2008. Egg size variation among tropical and
temperate songbirds: an embryonic temperature hypothesis. Proceedings of the National Academy of Sciences USA 105: 9268-9271.
Moreno, J. and J. L. Osorno. 2003. Avian egg color and sexual selection:
does eggshell pigmentation reflect female condition and genetic quality?
Ecology Letters 6: 803-806.
Moreno, J., J. L. Osorno, J. Morales, S. Merino, and G. Tomás. 2004. Egg
coloration and male parental effort in the Pied Flycatcher Ficedula hypoleuca. Journal of Avian Biology 35:300–304.
Pettingill, O.S., Jr. 1985.
Ornithology in Laboratory and Field, Fifth ed. Academic Press, New York, NY.
Pitcher, T.
E., P. O. Dunn & L. A. Whittingham. 2005. Sperm competition and the
evolution of testes size in birds. Journal of Evolutionary
Biology 18: 557-567.
Rosenzweig, M.R., A.L. Leiman and
S.M. Breedlove. 1996. Biological Psychology. Sinauer Associates, Sunderland, MA.
Rutstein, A.N., P. J. B.
Slater, and J. A. Graves. 2004. Diet
quality and resource allocation in the zebra finch. Proc. R. Soc.
Lond. B (Suppl.). Published online, 20 February 2004.
Saether, S. A., P. Fiske, &
J. A. Kalas. 2001. Male mate choice, sexual conflict and strategic allocation
of copulations in a lekking bird. Proceedings of the Royal Society London B 268: 2097 - 2102.
Samour, Jaime H. 2002. The
Reproductive Biology of the Budgerigar (Melopsittacus undulatus): Semen
Preservation Techniques and Artificial Insemination Procedures. Journal of
Avian Medicine and Surgery 16: 39-49.
Sato, T., Yen-nien Cheng,
Xiao-chun Wu, D. K. Zelenitsky, & Yu-fu Hsiao. 2005. A Pair of Shelled Eggs
Inside A Female Dinosaur. Science 308:375.
Siefferman, L., K. J. Navara, and
G. E. Hill. 2006. Egg coloration is correlated with female condition in Eastern
Bluebirds (Sialia sialis). Behavioral Ecology and Sociobiology 59:
651-656.
Sotherland, P. R. and H. Rahn.
1987. On the composition of bird eggs. Condor 89: 48-65.
van der Molen, W. H. 2002. Laying
an egg. http://www.afn.org/~poultry/egghen.htm.
Verboven, N., P. Monaghan, D.M.
Evans, H. Schwabl, N. Evans, C. Whitelaw, and R.G. Nager. 2003. Maternal
condition, yolk androgens and offspring performance: a supplemental feeding
experiment in the Lesser Black-backed Gull (Larus fuscus).
Proceedings of the Royal Society: Biological Sciences 270: 2223 - 2232
Visser, M. E., L. J. M. Holleman,
and S. P. Caro. 2009. Temperature
has a causal effect on avian timing of reproduction. Proceedings of the
Royal Society B: online early.
Welty, J.C. and L. Baptista.
1988. The life of birds, fourth ed. Saunders College Publishing, New York, NY.
Wingfield, J. C., and T. P. Hahn.
1994. Testosterone and territorial behaviour in sedentary and migratory
sparrows: Animal Behaviour 47: 77–89.
Wingfield, J. C., J. D. Jacobs,
A. D. Tramontin, N. Perfito, S. Meddle, D. L.
Maney, and K. Soma. 2000. Toward and ecological basis of hormone-behavior
interactions in reproduction of birds: In K. Wallen and J. Schneider (eds.),
Reproduction in context, pp. 85–128. MIT Press, Cambridge.
Winterbotton, M., T. Burke, &
T. R. Birkhead. 2001. The
phalloid organ, orgasm and sperm competition in a polygandrous bird: the
red-billed buffalo weaver. Behavioural Ecology and Sociobiology 50: 474-482.
Back to BIO 554/754
Syllabus
Back to Avian Biology










































{kind=link}
{kind=link}
{kind=link}
{kind=link}